
The Neuropharmacology of γ-hydroxybutyrate (GHB)
Related to Psychoactive Issues
v2 Jan 2004 (first revision June 2003)
edited & published by Erowid
Citation: BilZ0r. "Neuropharmacology of Gamma-HydroxyButyrate".
Erowid.org, v2 Jan 2004 (v1 June 2003): chemicals/ghb/ghb_pharmacology1.shtml.
γ-Hydroxybutyrate (GHB) is a central nervous system depressant. It has been used as an anxiolytic, anaesthetic and sedative/hypnotic4, inducing a sleep-like state in experimental animals in doses ranging from 0.1 to 1.5g/kg9,5,11. GHB is also a putative neurotransmitter4. Treatment with GHB has been shown to affect many neurotransmitter systems, most notably, dopamine, glutamate and acetylcholine4. A number of early studies focused on GHB's action at dopaminergic synapses and showed that GHB initially inhibits dopamine release and then, time-dependently, causes its release6. This effect has now been shown to be due to experimental errors, such as ignoring the effects of anaesthetics, or having unusually high levels of Ca2+ in the growth medium6.
The mechanism of how GHB effects the CNS has been unclear up until now. Resolving the mechanism of GHB's action has been complicated by the fact that GHB acts on both the GABA-B and the newly sequenced GHB receptor4. The evidence that GHB affects the GABA-B receptor is threefold. On a behavioral level, antagonizing the GABA-B receptor stops the discriminative effects of GHB3. On a cellular level, GHB causes a pronounced hyperpolarization of nerve cells and this effect is blocked by GABA-B antagonists but not by GHB receptor antagonists13. Finally, on a molecular level GHB binds (although weakly, KD =100�M10) to the GABA-B receptor.
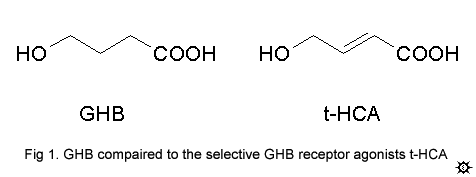
The GHB receptor is a novel G-protein coupled receptor1, which is widely distributed throughout the cortex and certain subcortical locations (most notably the hippocampus, amygdala, septum, basal ganglia and substantia nigra) (Hechler et al., 1992). Importantly, GHB exhibits an extremely high affinity for the GHB receptor2,12. It is obvious that GHB receptors mediate some of GHB's action because cortical neurons which are normally activated by GHB revert to their normal activity levels when GHB is added, along with its selective receptor antagonist NCS-3827. The recent development of truly selective GHB receptor agonists has allowed scientists to finally understand GHB's mechanism. At low doses, GHB is binding almost exclusively to the GHB receptor, a receptor which, through pathways not yet understood, stimulates the release of glutamate (the major excitatory neurotransmitter) in the cortex. As the dose of GHB increases, more and more GHB is binding to, and activating, the inhibitory GABA-B receptor. This leads to the sedative/hypnotic effect of GHB. The selective GHB receptor agonist t-HCA (fig 1) does not cause sedation of any kind, and is presumably a stimulant as it leads to seizures4.
These findings explain many things about GHB research and the subjective experience brought on by GHB. For instance, the reason why some experiments would find the GABA-B receptor to be the main mediator of GHB's effects was because they used high doses, or were measuring effects produced by the GABA-B receptor. On the other hand, experimenters using lower doses or measuring cortical excitation found the GHB receptor to be the main mediator. This also explains why people who take GHB to sleep would wake up 2-3 hours after dosing. After the initial dosing, the GABA-B receptor was being stimulated, leading to sleep, but as the GHB was being metabolised and the concentration in the body dropped, the GABA-B receptor stopped being activated, and the excititory GHB receptor became the main receptor being activated.
The mechanism of how GHB effects the CNS has been unclear up until now. Resolving the mechanism of GHB's action has been complicated by the fact that GHB acts on both the GABA-B and the newly sequenced GHB receptor4. The evidence that GHB affects the GABA-B receptor is threefold. On a behavioral level, antagonizing the GABA-B receptor stops the discriminative effects of GHB3. On a cellular level, GHB causes a pronounced hyperpolarization of nerve cells and this effect is blocked by GABA-B antagonists but not by GHB receptor antagonists13. Finally, on a molecular level GHB binds (although weakly, KD =100�M10) to the GABA-B receptor.
The GHB receptor is a novel G-protein coupled receptor1, which is widely distributed throughout the cortex and certain subcortical locations (most notably the hippocampus, amygdala, septum, basal ganglia and substantia nigra) (Hechler et al., 1992). Importantly, GHB exhibits an extremely high affinity for the GHB receptor2,12. It is obvious that GHB receptors mediate some of GHB's action because cortical neurons which are normally activated by GHB revert to their normal activity levels when GHB is added, along with its selective receptor antagonist NCS-3827. The recent development of truly selective GHB receptor agonists has allowed scientists to finally understand GHB's mechanism. At low doses, GHB is binding almost exclusively to the GHB receptor, a receptor which, through pathways not yet understood, stimulates the release of glutamate (the major excitatory neurotransmitter) in the cortex. As the dose of GHB increases, more and more GHB is binding to, and activating, the inhibitory GABA-B receptor. This leads to the sedative/hypnotic effect of GHB. The selective GHB receptor agonist t-HCA (fig 1) does not cause sedation of any kind, and is presumably a stimulant as it leads to seizures4.
These findings explain many things about GHB research and the subjective experience brought on by GHB. For instance, the reason why some experiments would find the GABA-B receptor to be the main mediator of GHB's effects was because they used high doses, or were measuring effects produced by the GABA-B receptor. On the other hand, experimenters using lower doses or measuring cortical excitation found the GHB receptor to be the main mediator. This also explains why people who take GHB to sleep would wake up 2-3 hours after dosing. After the initial dosing, the GABA-B receptor was being stimulated, leading to sleep, but as the GHB was being metabolised and the concentration in the body dropped, the GABA-B receptor stopped being activated, and the excititory GHB receptor became the main receptor being activated.
References #
- Andriamampandry C, Taleb O, Viry S, Muller C, Humbert JP, Gobaille S, Aunis D, Maitre M. Cloning and characterization of a rat brain receptor that binds the endogenous neuromodulator gamma hydroxybutyrate (GHB). Faseb J. 17(12):1691-3, 2003.
- Benavides J, Rumigny JF, Bourguignon JJ, Cash C, Wermuth CG, Mandel P, Vincendon G, Maitre M. High affinity binding sites for gamma-hydroxybutyric acid in rat brain. Life Sci. 15;30(11), 953-61, 1982.
- Carter LP, Flores LR, Wu H, Chen W, Unzeitig AW, Coop A, France CP. The Role of GABAB Receptors in the Discriminative Stimulus Effects of gamma -Hydroxybutyrate in Rats: Time Course and Antagonism Studies. J Pharmacol Exp Ther. 2003. 305(2), 668-74.
- Castelli MP, Ferraro L, Mocci I, Carta F, Carai MA, Antonelli T, Tanganelli S, Cignarella G, Gessa GL. Selective gamma-hydroxybutyric acid receptor ligands increase extracellular glutamate in the hippocampus, but fail to activate G protein and to produce the sedative/hypnotic effect of gamma-hydroxybutyric acid. J Neurochem. 2003 Nov. 87(3):722-32.
- Drakontides AB, Schneider JA, Funderburk WH. Some effects of sodium gamma-hydroxybutyrate on the central nervous sytem. J Pharmacol Exp Ther. 135, 275-84, 1962.
- Feigenbaum JJ, Howard SG. Does gamma-hydroxybutyrate inhibit or stimulate central DA release? Int J Neurosci. 88(1-2), 53-69, 1996.
- Godbout R, Jelenic P, Labrie C, Schmitt M, Bourguignon JJ. Effect of gamma-hydroxybutyrate and its antagonist NCS-382 on spontaneous cell firing in the prefrontal cortex of the rat. Brain Res. 27;673(1), 157-60, 1995.
- Hechler V, Gobaille S, Maitre M. Selective distribution pattern of gamma-hydroxybutyrate receptors in the rat forebrain and midbrain as revealed by quantitative autoradiography. Brain Res. 1992. 572(1-2):345-8.
- Laborit H. Sodium 4-hydroxybutyrate. Int J Neuropharmacol. 3, 433-52, 1964.
- Mathivet P, Bernasconi R. Binding characteristics of gamma-hydroxybutyric acid as a weak but selective GABA B receptor agonist. Eur J Pharmacol. 321(1), 67-75, 1997.
- Tunnicliff G. Significance of γ-hydroxybutyric acid in the brain. Gen Pharmacol. 23, 1027-34, 1992.
- Tunnicliff G. Sites of action of gamma-hydroxybutyrate (GHB) - a neuroactive drug with abuse potential. J Toxicol Clin Toxicol. 35, 581-90, 1997.
- Williams SR, Turner JP, Crunelli V. Gamma-hydroxybutyrate promotes oscillatory activity of rat and cat thalamocortical neurons by a tonic GABAB, receptor-mediated hyperpolarization. Neuroscience. 66(1), 133-41.
Additional References #
- Anden NE, Magnusson T, Stock G. Effects of drugs influencing monoamine mechanisms on the increase in brain dopamine produced by axotomy or treatment with gamma-hydroxybutyric acid. Naunyn-Schmiedebergs Arch Pharmacol. 26;278(4), 363-72, 1973.
- Cook CD, Aceto MD, Coop A, Beardlsey PM. Effects of the putative antagonist NCS382 on the behavioral pharmacological actions of γ-hydroxybutyrate in mice. Psychopharmacology (Berl). 160(1), 99-106, 2002.
- Hu RQ, Banerjee PK, Snead OC 3rd. Regulation of gamma-aminobutyric acid (GABA) release in cerebral cortex in the gamma-hydroxybutyric acid (GHB) model of absence seizures in rat. Neuropharmacology. 28;39(3), 427-39, 2000.
- Lingenhoehl K, Brom R, Heid J, Beck P, Froestl W, Kaupmann K, Bettler B, Mosbacher J. Gamma-hydroxybutyrate is a weak agonist at recombinant GABA(B) receptors. Neuropharmacology. 38(11), 1667-73, 1999.
- Maitre M. The gamma-hydroxybutyrate signalling system in brain: organization and functional implications. Prog Neurobiol. 51(3), 337-61, 1997.
- Maitre M, Hechler V, Vayer P, Gobaille S, Cash CD, Schmitt M, Bourguignon JJ. A specific gamma-hydroxybutyrate receptor ligand possesses both antagonistic and anticonvulsant properties. J Pharmacol Exp Ther. 255(2), 657-63, 1990.
- Snead OC 3rd. Evidence for a G protein-coupled gamma-hydroxybutyric acid receptor. J Neurochem. 75(5), 1986-96, 2000.
- Walters JR, Roth RH, Aghajanian GK. Dopaminergic neurons: similar biochemical and histochemical effects of gamma-hydroxybutyrate and acute lesions of the nigro-neostriatal pathway. J Pharmacol Exp Ther. 186, 630-9, 1973.
- Gobaille S, Hechler V, Andriamampandry C, Kemmel V, Maitre M. γ-Hydroxybutyrate modulates synthesis and extracellular concentration of gamma-aminobutyric acid in discrete rat brain regions in vivo. J Pharmacol Exp Ther. 290(1), 303-9, 1999.